The Ediacaran is a Precambrian (Neoproterozoic) assemblage, which existed from about 610 to 560 million years ago. The fauna has been found on all continents except Antarctica, with the most important sites in Newfoundland, Canada, Namibia, the White Sea Coast, Russia and the Flinders Ranges South Australia (Jensen et al., 1998).
One of the best localities where the significance of the fauna was first recognised was in South Australia (Sprigg, 1947, Figure 1). The name Ediacara comes from the site where the fossils were first found. Whilst the fauna has a world-wide distribution, it is important to note that there are significant differences in the diversity of the fauna at different localities, due primarily to different environmental conditions. The Australian and Russian forms are similar and the rocks they are preserved in are indicative of a shallow water ecosystem. The Newfoundland forms are markedly different and the rocks are indicative of a deep water setting (Jensen et al., 1998).
Figure 1. Outcrops of the Rawnsley Quartzite.
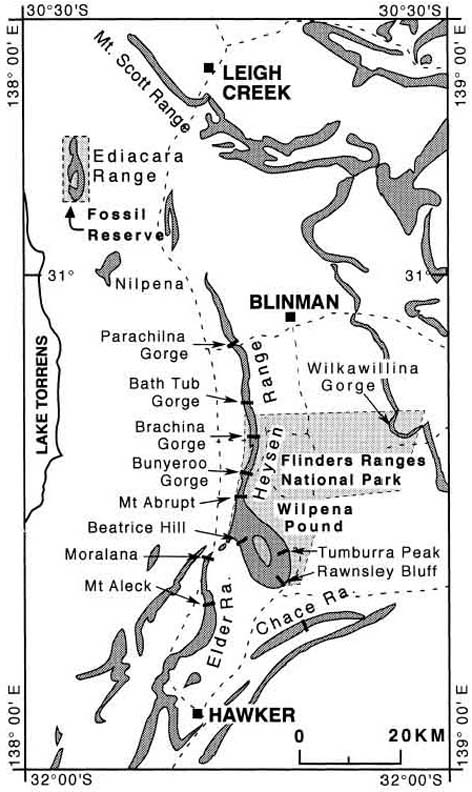
Locality map for outcrop locations of the Rawnsley Quartzite, Pound Subgroup in the Flinders Ranges, South Australia. The National Park and fossil reserve are shaded. From Gehling (2000).
In South Australia, the fossils occur as depressions up into or extensions down from the bottom of thin quartzite beds (Glaessner and Wade, 1966). The fossils occur in preservational windows in the Ediacara Member of the Rawnsley Quartzite, a formation of the Pound Subgroup, bounded above by the Early Cambrian Uratanna sequence (Gehling, 2000). The Rawnsley depositional sequence is developed over an erosional surface having approximately 250 m of relief, with southeasterly paleovalleys are filled with sequences of massive sandstone and laminated siltstones, grading up into wellbedded sandstone, Figure 2. The fossils occur above the valley fill facies, on sandstone partings within upward-shoaling, delta-front environments between storm- and fairweather wave base (Gehling et al., 2000). The clay lenses were subsequently highly compacted and altered and are now mostly only thin, lenticular partings between the quartzite flags. Most of these partings can be opened only by natural weathering. The Enorama Creek section of Flinders was designated the "boundary stratotype" for the Ediacaran by the Terminal Proterozoic Period subcommission. A boundary stratotype is a rock sequence and level that is defined and used as the standard comparison for all other rock sequences of its age.
Figure 2. Generalised section through the Rawnsley Quartzite.
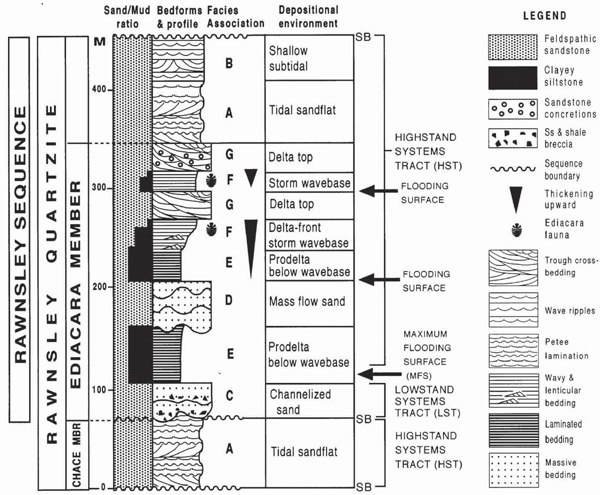
Generalised section including depositional environments of the Rawnsley Quartzite, Flinders Ranges, South Australia. From Gehling (2000).
Ediacaran fauna are a diverse group with more than 30 genera named, however, they can be categorised into four broad morphological categories. Most abundant are circular impressions, some of which appear similar to medusoid cnidarians (Gehling et al.,2000). Others of this form are believed to be the holdfasts of the frond-like Ediacarans. There are also trace fossils of various tracks and burrows made by bilaterian animals (Jenkins et al., 1983). The third most abundant fossils are a number of benthic forms, some of these suggest affinities with extant groups such as annelids (e.g. Dickinsonia and Spriggina, echinoderms (e.g. Arkarua) (Gehling, 1987), or arthropods (e.g. Diplichnites and Parvancorina). Least abundant, though perhaps most characteristic of the assemblage as a whole, are the attached, frond-like organisms which some propose have affinities with the sea pens and other soft corals. A typical Ediacaran species had a soft body, yet they most commonly occur in silt- and sandstones which typically form in quite turbulent conditions: not the sort of sediments where soft tissue preservation occurs (Jenkins et al., 1983). Interestingly, it appears that Ediacaran communities were largely free of predators; no species appears to have possessed a jaw apparatus suitable for seizing and tearing prey, and few fossils show clear evidence of predatory damage.
The dominant factors which allowed preservation of the Ediacara fauna were: collagen, no bioturbation, and no predation. The ability to produce collagen was important because collagen is relatively inert, strong and flexible. A collagen outer layer helped hold the organism together and also allowed the fossil to retain shape when covered by the mantling sand. Also, since collagen was a relatively new compound (synthesis probably related to a threshold level of oxygen in the atmosphere), there was a lag before the micro-community used collagen as a food source (Narbonne and Gehling, 2003). The lower oxygen levels meant that bodies had to be kept small or thin, because oxygen was absorbed through the surface of the organism. Therefore, tissues had to be close to the surface in order to obtain oxygen by simple diffusion. There is little constraint to the size such organisms could reach, provided they stayed thin, hence half metre long 'flat' worms (Glaessner, 1958). Flat, thin bodies are poorly developed for burrowing, which meant that any organism that was buried was not disturbed by bioturbation. However, these conditions did not last. An interrelated series of events that included a rise in oxygen levels, the acquisition of mineralisation capabilities, the rise of predation and the ability to produce a round cross-sectional body-plan (oxygen depended) conducive to burrowing, soon demolished what was a pristine preservational environment.